幼苗发育是受光控制的。播种在土中的禾谷类作物种子,芽鞘和中胚轴在土中不断伸长,直到见光为止。而幼苗见光后,卷曲的叶片才张开
光可以抑制多种作物根的生长,光强度与抑制根生长呈正相关
虽然禾本科植物的叶片一旦冲破胚芽鞘,在一段时间内,在黑暗中生长一般在光下一样快,但在缺光时,双子叶植物叶片不能正常发育
光促进叶片扩大,主要是由于加强细胞分裂,细胞最终的大小和保持在黑暗中的并没有明显不同
在完整叶片中,亮光下生长的叶片的细胞的分裂伸长的分化速度比在低强度光下更快
对于在日光下发芽的植物,高强度的蓝光对茎伸长的抑制作用最大,而和它大约相等强度的红光促进伸长
在黑暗中发芽时蓝光作用较弱,绿光几乎无效。这说明光对黄花苗和绿苗的影响是不同的
在能量低时,红光比蓝光更具抑制作用,但随着强度的增加,蓝光变得更有效
当种子吸涨后,他们的发芽常受到光的影响。大约有一半的种子需要光照才发芽最好,也有许多种子的发芽不受光的影响,而有一些种子是受光抑制的
植物营养生长到生殖生长的转变是由红光-远红光受体和蓝光-近紫外光受体调节的
植物开花有光周期现象,日照长度对于植物从营养生长到花原茎形成这段时间的长短有决定性影响
研究表明,植物开花的光周期现象,在光期和暗期中,对于诱发花原茎形成起决定作用的是暗期长短
短日照植物必须在超过某一临界暗期时才能形成花芽
长日照植物必须在短于某一临界暗期时才能开花
闪光试验证明了暗期的重要性
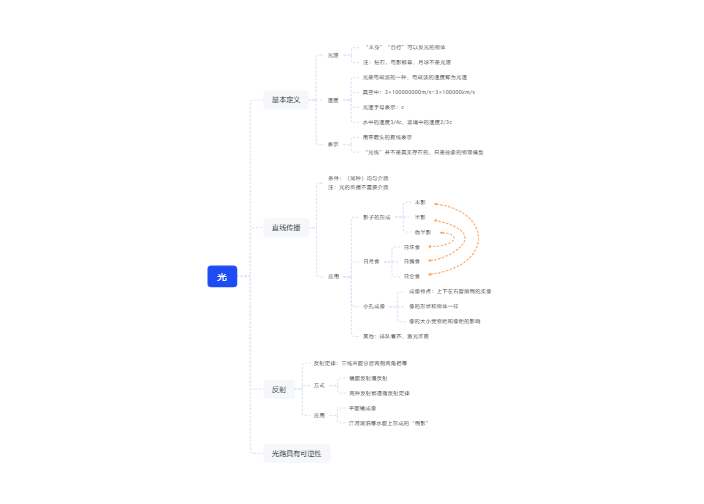
向光性:植物随光的方向而弯曲 如叶子能尽量伸展于最适宜利用光能的位置
产生向光性的原因:单侧方向的光照,可引起生长素向背光侧的横向运输,从而使背光侧的生长素多,细胞伸长强烈,所以植株便向光生长
对向光性起作用的是短波光,红光无效
许多研究表明,温带植物的秋季落叶、冬季休眠与日照长度有关
如杨树在给予几天短日照后,即使气温还在相当高的情况下,但在继续长出几片叶子后即形成顶芽,叶子的生长停止
如再继续短日照处理(温度条件不变),则其叶子便不再生长,然后逐渐萎黄脱落,进入深休眠
如果再给以长日照处理,则植物可继续生长,而不进入休眠状态。
洋槐、柳等树种对短日照也有同样的反应。因此北方植物园在引种工作中,可利用短日照促使树木提早休眠,增强御寒越冬的能力
叶绿素的合成离不开光的参与。叶绿素的合成与光时和光质有关。不同光质下合成的叶绿素a和叶绿素b的比例有所差别
红光更有利于叶绿素a的形成
黄光和蓝光更有利于叶绿素b的形成
光促进花青素苷和其他类黄酮化合物形成
植物在生命活动过程中所产生的全部有机物质的碳骨架都来自于光合作用,而光合作用的能量来源是可见光
光敏色素有两种类型:红光吸收型Pr和远红光吸收型Pfr
当Pr吸收660nm红光后,就转变为Pfr,而Pfr吸收730nm远红光后,会逆转为Pr
光敏色素在植物生命活动中作用很广泛
光敏色素可以影响质体的形成,原生质体膨大,膜透性的改变,细胞的分化及花色素的形成
一些需光种子如莴苣、烟草和拟南芥等植物的种子萌发时需要光照,在红光下萌发率高,在远红光下萌发率低。
另外光敏色素在光形态建成中的作用还表现在根原基起始,叶分化与扩大,弯钩张开,子叶张开,单子叶植物叶片开展,叶片偏上生长,节间延长,叶片肉之花等方面
子叶运动,含羞草小叶运动,节律现象,叶脱落,休眠,储藏器官的形成和生长
引起光周期反应,诱导花芽分化开花,性别表现,花粉育性
调控内源激素代谢及运输
如磷酸甘油醛脱氢酶、转酮醇酶、RNA聚合酶、核糖核酸酶、苯丙氨酸裂解酶
调节Rubisco的基因表达
引起蓝光反应,调节高等植物的向光反应、叶绿体的向光性反应、开花的光周期
抑制茎的伸长、促进气孔开放、幼苗去黄化反应
生理钟、促进花色素苷积累及调节基因表达等生理过程
蓝光受体有隐花色素和向光素,可以调节不同的蓝光反应
隐花色素参与调节抑制茎的伸长、幼苗去黄化反应、开花的光周期、生理钟以花色素苷合成酶基因的表达
向光素主要调节植物的运动,如向光反应、气孔运动及叶绿体运动等
UV-B紫外光可引起类黄酮、花色素苷等色素合成的增加
例如能降低黄瓜、小麦、大豆、玉米、水稻等的株高,减少叶面积和光合能力。
因此在表皮细胞中由紫外线诱导形成的、能够吸收UV-B的花青素苷和黄酮类物质,可能是植物的一种自我保护反应
主要是蓝紫光,尤其是紫外光
光对植物的高矮、株型、分枝的多少、长度、根冠比、叶片的大小颜色及生长特性等形态建成的直接作用通常与光敏色素有关。例如黄化苗转绿
通常促进需光种子的萌发,远红光抑制需光种子的萌发
绝大多数多年生植物都是长日照条件下促进生长,短日照条件下诱导休眠
光的方向对生长有影响,通常茎叶有正向光性,根有负向光性
植物的光周期与成花诱导
光是光合作用的能源,并且光是叶绿素形成的条件
光合产物是生长物质的基础
参与高能反应