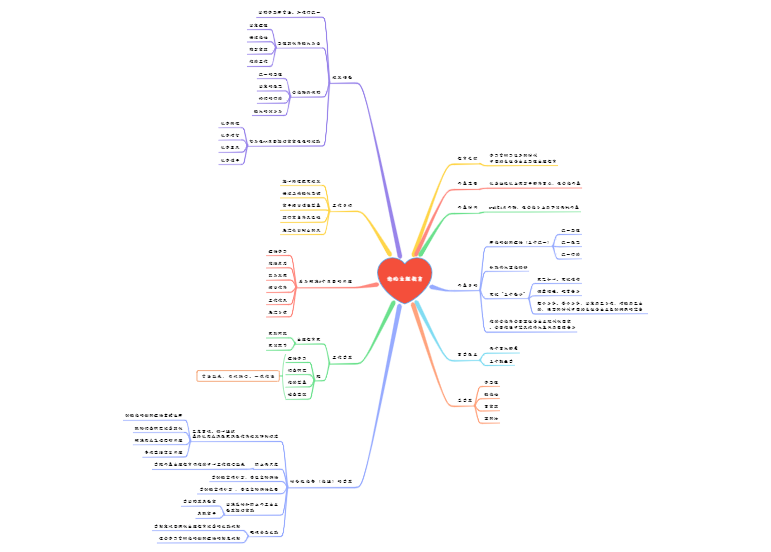
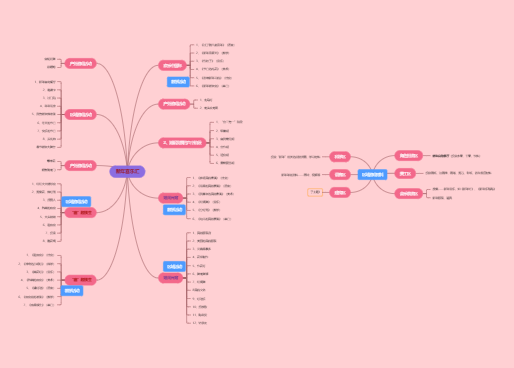
能够加工抗原并以抗原肽-MHC分子复合物的形式将抗原肽提呈给T细胞
一类被胞内病原体感染而产生病原体抗原或细胞发生突变产生突变蛋白抗原的细胞
树突状细胞(DC)、单核/巨噬细胞(Mo、MØ)、B细胞
组成性表达MHC-Ⅱ类分子、共刺激分子、黏附分子
直接摄取、加工和提呈抗原
内皮细胞、上皮细胞、成纤维细胞
通常不表达或低表达MHC-Ⅱ类分子
加工、提呈抗原能力弱
在炎症或某些IFN-ɣ(CK作用)等诱导下表达MHC-Ⅱ类分子、共刺激分子、黏附分子
如靶细胞、内皮细胞、上皮细胞、激活的T细胞等
一类成熟时具有许多树突样突起、能够识别摄取加工外源性抗原并将抗原肽提成给初始T细胞诱导其活化增殖的功能最强的APC
表达参与抗原社区和转运的特殊膜受体
是抗原提呈能力最强的APC
能活化初始T细胞(唯一)
作用:主要参与适应性免疫应答的诱导和启动
分布:各种实体组织和上皮细胞
未成熟DC在趋化因子作用下,由外周组织器官进入淋巴管或血液循环至外周淋巴管器官的处于迁移阶段(不成熟)的DC
未成熟DC→接触摄取抗原或某些炎症刺激→表达特定的趋化因子受体(如CCR7)
特点:表面有许多树突样突起;低表达PRR——识别摄取外源性抗原能力弱;加工抗原能力弱;高表达MHC-Ⅱ类分子、共刺激分子、黏附分子
并指状DC(IDC)——外周免疫器官T细胞区的成熟DC
黏膜—成熟DC—诱导黏膜局部的免疫应答
胸腺—胸腺DC—诱导T细胞中的中枢免疫耐受
特殊——调节性DC:具有负向调控免疫应答维持免疫耐受的作用
作用:活化后快速产生大量IFN-Ⅰ→参与抗病毒固有免疫应答;某些情况下参与自身免疫病的发生与发展
特点:树突状,不具备抗原提呈能力
作用:负载抗原肽刺激生发中心B细胞发生体细胞超突变
表达PRR,识别PAMP 如MR、Toll样受体
表达Fc-R
识别、摄取、杀灭
意义:DC最重要的功能(唯一激活初始T细胞的APC)
成熟DC→CD80/40/86等共刺激分子→T细胞活化的第二信号
Ag→DC→抗原肽-MHC-Ⅱ类分子复合物→CD4+ T细胞→Th0细胞活化的启动信号
DC→CK→诱导活化T细胞增殖分化并进行免疫应答
DC→高表达ICAM-1等 CAM→促进细胞间的相互作用
Ag→DC抗原肽-MHC-Ⅰ类分子复合物→CD8+ T细胞
DC→诱导Ig的类别转换、促进B细胞增殖分化、参与体液免疫
机制:CK、趋化因子→调节免疫细胞的分化发育、活化、移行等
经典:DC→ IL-2 →Th0→ Th1 →产生Th1型免疫应答
未成熟DC:参与外周免疫耐受的诱导
胸腺DC:T细胞阴性选择,中枢免疫耐受
专职性APC
BCR识别、浓集和内化抗原
抗原肽-MHCⅡ类分子复合物→Th
在抗原浓度极低时仍能提呈抗原(浓集抗原的作用);激活Th的同时,B细胞也受Th辅助活化
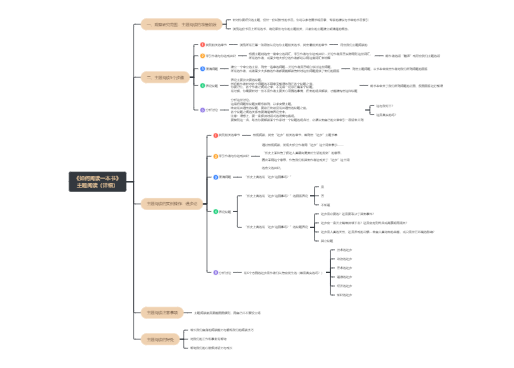
APC将摄取入胞内的外源性抗原或者胞质内产生的内源性抗原降解,使抗原肽与MHC结合形成复合物转运到细胞表面的过程
表达于APC表面的抗原肽-MHC分子复合物被T细胞识别、诱导T细胞活化
外源性:如被吞噬的细胞、细菌或蛋白抗原等,来自细胞外的抗原
内源性:如病毒感染细胞内合成的病毒蛋白、肿瘤细胞内合成的肿瘤抗原和某些细胞内的自身抗原等
泛素化-蛋白酶体和免疫蛋白酶体→内源性抗原→抗原肽
TAP1+2,转运8-12aa的抗原肽到内质网(ER)
特点
MHC-Ⅰ——ɑ链和β2m在ER合成
ER内的氨基肽酶——ERAP
羟基氧化还原酶Erp57
复合物→高尔基体→质膜→CD8+ T细胞
实质:外源性/溶酶体抗原提呈途径
机制
特点:MHC-Ⅱ类分子的抗原结合槽两端开放,13~17个aa
MⅡC内,HLA-DM介导抗原肽结合槽与CLIP的解离和与抗原肽的结合(复合物)
复合物→质膜→CD4+ T细胞
特点:部分抗原肽可不经过li依赖途径直接与质膜表面空载的MHC-Ⅱ类分子结合后提呈
MHC分子对抗原的交叉提呈/致敏途径,使参与机体针对病毒、细菌和大多数肿瘤的免疫应答。不是抗原提呈的主要方式,也不涉及MHC分子的合成
外源性
内源性
机制(见书)
CD1d——NK细胞,参与固有免疫应答
CD1a~c——T细胞,介导适应性免疫应答