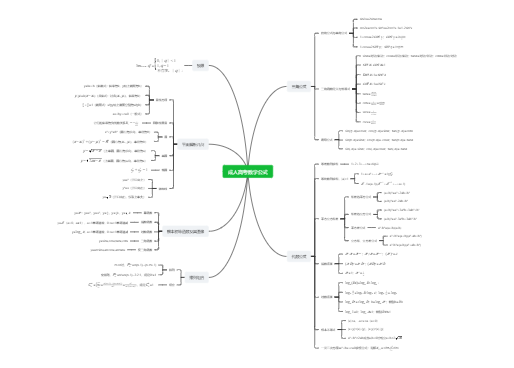
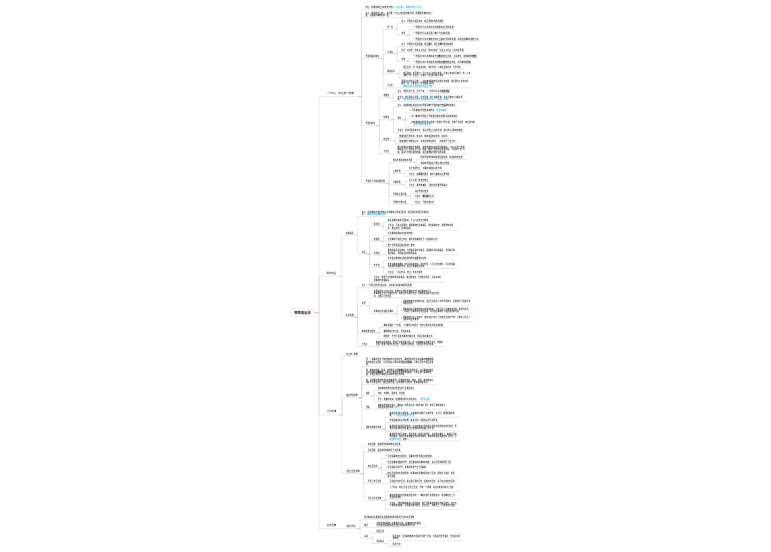
3个13bp串联重复序列(连续)
4个9bp保守序列(分散)
DNA解链酶(水解ATP获得能量)eg:DnaB
1.大多数-->沿后随链模板的5'->3'方向并随复制叉前进而移动
2.Rep蛋白-->沿前导链模板3'->5'方向移动
推测Rep蛋白和特定DNA解链酶分别在DNA两条母链上协同作用,
==>解开DNA双链
SSB (single-strand binding protein),单链结合蛋白
保证被解链酶解开的单链在复制完成前保持单链结构,
待单链复制完成后离开,重新进入循环
拓扑异构酶
消除解链造成的正超螺旋的堆积,消除阻碍解链继续进行的压力
==>解链得以继续,复制得以延伸
前导链只需一次引发,而滞后链需要多次引发
引发前体(6种蛋白质,DnaB、DnaC等)+引发酶(DnaG)
特异的RNA聚合酶,仅用于合成复制所需的一小段RNA引物
作用过程
1.ATP作用下,DnaA蛋白与oriC的4个9碱基保守序列结合,形成寡聚复合物
2.在DNA结合蛋白HU蛋白和ATP共同作用下,DnaA复制起始复合物使3个13碱基串联重复序列变性,形成开链
3.在DnaC蛋白协助下,解链酶DnaB替换DnaA与DNA链结合,使DNA双链解开
4.引物合成(以后随链为例)
引发体在后随链上前进,在模板上断断续续地引发生成后随链的引物RNA短链;
【再由DNA聚合酶Ⅲ作用生成DNA,直至遇到下一个引物或冈崎片段为止;
由RNase H降解RNA引物并由DNA聚合酶Ⅰ将缺口补齐;
再由DNA连接酶将冈崎片段连接形成大分子DNA】
模板、引物【RNA、DNA、Pr.】、dNTP、3'-OH
1.Klenow(大)--具DNA聚合酶活性、3'-->5'核酸外切酶活性
2.小片段--具5'-->3'核酸外切酶活性
切除引物,填补缺口(gap≠nick)
具5'-->3'核酸外切酶活性,起修复作用
具5'-->3'聚合作用,主要的复制酶
全酶异二聚体
连接切除引物后留下的切口(nick)
DNA聚合酶全酶异二聚体中的一个亚单位与前导链模板结合,
另一个亚单位与后随链模板结合
DNA聚合酶Ⅲ以5'-->3'方向延伸引物,合成子链
DNA聚合酶Ⅰ具5'-->3'外切酶活性,切除引物
DNA聚合酶Ⅲ延伸,直到遇见邻近引物的5'端
DNA聚合酶Ⅰ切除引物
DNA连接酶连接相邻冈崎片段,形成完整子链
复制叉前移遇到重复终止子序列(Ter)时,Ter-Tus复合物使DnaB不再解链,阻挡复制叉的前进,等到相反方向的复制叉到达后停止复制,拓扑异构酶作用下释放子链
引物合成,无校对作用(大肠杆菌的DnaG)
损伤修复(大肠杆菌的聚合酶Ⅱ)
线粒体DNA复制
主要的DNA复制酶(大肠杆菌的聚合酶Ⅲ),与PCNA(增殖细胞核抗原)结合
复制修复,相当于大肠杆菌的聚合酶Ⅰ
酵母的复制起始点称为自主复制序列(ARS)
其他真核生物中也存在类似酵母ARS元件的序列
不同ARS序列都具有富含AT碱基对的A区
真核生物DNA复制起始需要起始点识别复合物(ORC)参与
复制叉上存在引发酶复合体(DNA聚合酶α)和两种DNA聚合酶复合体:
DNA聚合酶δ负责延伸前导链;DNA聚合酶ε负责合成后随链的冈崎片段
可特异性切除DNA-RNA杂合底物的RNA酶H1发挥核酸内切酶活性切开引物
由具5'->3'外切酶活性的特异蛋白降解RNA片段
由DNA连接酶Ⅰ连接相邻冈崎片段
DNA复制的调控主要发生在起始阶段,一旦开始复制,
若无意外阻力,则将一直复制直到完成
起始物位点编码的复制调节蛋白质可以与复制复合物相互作用,
确定复制起始频率和复制方式
又称限制点调控,决定细胞停留在G1期还是进入S期
许多外部因素(促细胞分裂素、致癌剂、外科切除)参与限制点调控
决定不同染色体或同一染色体不同部位的复制子按一定顺序在S期起始复制
决定复制的起始与否