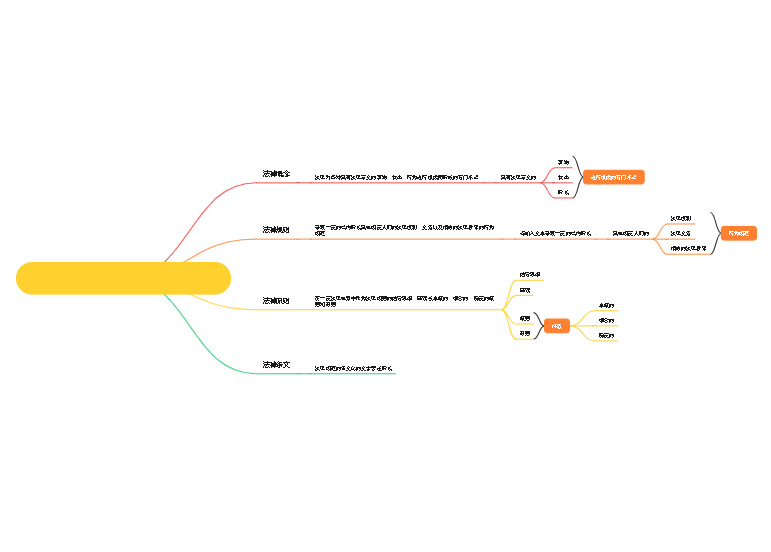
单核苷酸
高等动物体内的干扰素可诱导靶细胞合成一种依靠此键连接的多聚腺苷酸
区分RNA和DNA的唯一标准
会亲核进攻旁边的3',5'-磷酸二酯键,产生2',3'-磷酸二酯键
适合mRNA
2'-OH变成O-,亲核性大大增加
RNA一般在酸性条件下存放
tRNA、rRNA等可能将2'-OH甲基化以增强自身稳定性
利用羟基的亲核进攻性,作为酶催化一些重要的生化反应
RNA中的T来自U的甲基化
细胞中存在少量dUTP,在DNA复制时可直接参入
导致突变的产生
将U切除
根据对面碱基合成新的碱基
防止C脱氨基产生的“坏U”和本来的“好U”混淆
密码子的简并性
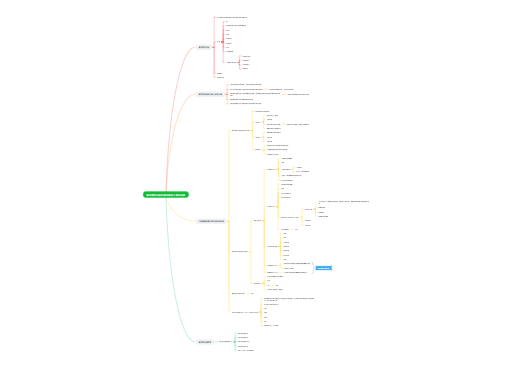
RNA不是遗传物质
RNA比DNA小得多,发生自发脱氨基的概率小得多
同一种RNA有大量拷贝,个别突变不影响整体功能
细胞内有一套质量控制系统,能即时水解错误的RNA
使RNA能折叠成各种三维结构
易于复制
易于修复
核小RNA
仅真核生物
帮助确定外显子、内含子的交界
作为酶催化剪接反应
核仁小RNA
真核生物和古菌
通过碱基互补配对定位rRNA上需要修饰的碱基
干扰RNA
特异性干扰翻译过程,控制基因表达
微RNA
细胞内源
仅真核生物
小干扰RNA
细胞外源
转移信使RNA
仅细菌
参与细菌无终止密码子的mRNA的抢救翻译
仅雌性哺乳动物表达
促进哺乳动物一条X染色体转变成高度浓缩的巴氏小体
编码RNA
呈组成型表达,是细胞的正常功能和生存所必须的
只在特定的细胞,或在生物发育的某个阶段,或受到特定的外界刺激后才表达
其表达能够在转录或翻译水平上影响到其他基因的表达
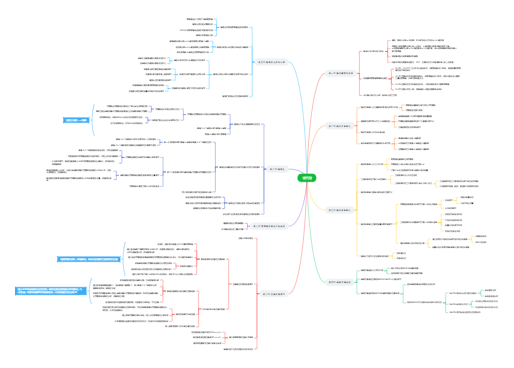
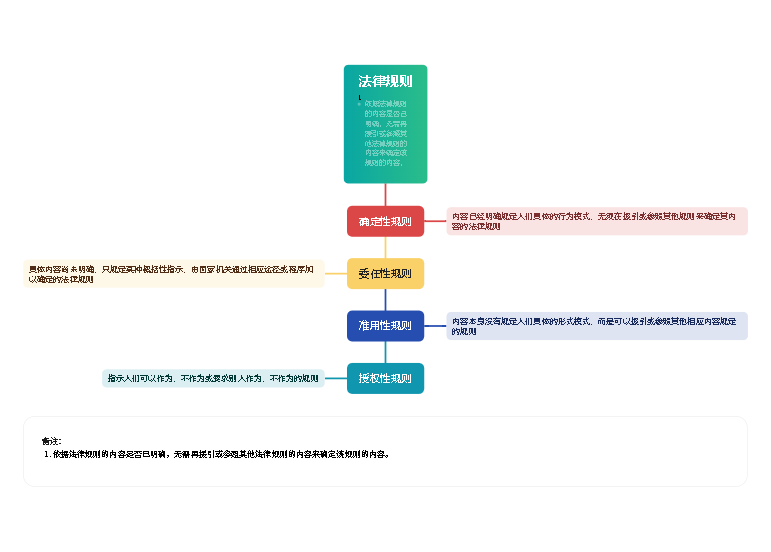
构成核酸的多聚核苷酸链上所有核苷酸或碱基的排列顺序
称为极性
在生理pH下,多聚核苷酸链的磷酸基团大多处于解离的状态,带有大量负电荷
每一条链上的核苷酸残基都有一定的排列顺序
线形
细菌的染色体DNA
大多数质粒DNA
叶绿体DNA
大多数线粒体DNA
5'→3'
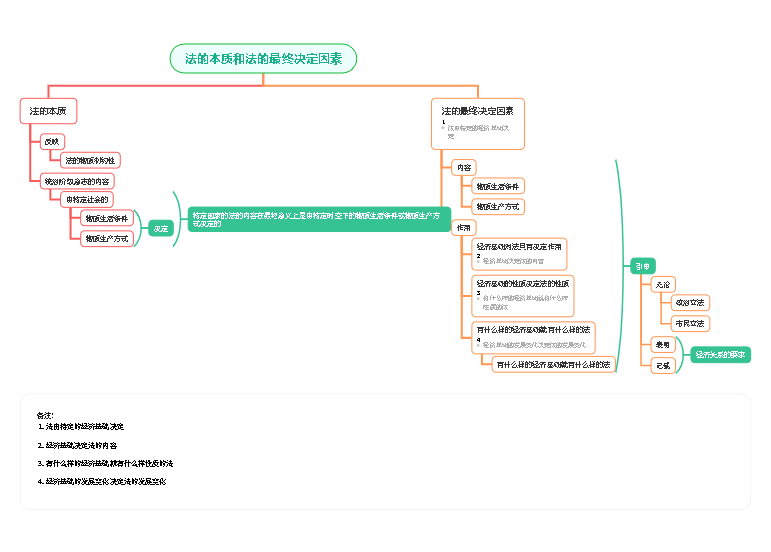
生物体的遗传信息是贮存在有4种核苷酸编码的特定的序列之中,而与高级结构无关
相对湿度
盐的种类和浓度
碱基组成和序列
超螺旋的数量和方向
由Watson和Crick提出
DNA由两条反平行的多聚脱氧核苷酸链组成,两条链互相缠绕,形成右手双螺旋
最常见的双螺旋
更重要
可以用于与其他生物分子发生特异性相互作用
碱基对位于双螺旋的内部,并垂直于螺旋轴,而磷酸脱氧核糖骨架位于螺旋表面
螺旋-环-螺旋
螺旋-转角-螺旋
一个α螺旋深入大沟,与其中剩余的氢键供受体作用
大沟内碱基对暴露得非常清楚,可以区分GC、CG
0.33nm
36°
2.00nm
一圈螺旋的高度
3.32nm
10bp
细胞内实际约10.4-10.6bp
相对湿度(含水量)的降低
盐浓度的上升
11bp
2.46nm
2.6nm
19°
0.23nm
窄而深
与A型双螺旋作用的蛋白质通常作用于小沟(包括与RNA作用)
某些革兰氏阳性菌孢子内的DNA
DNA复制时与DNA聚合酶活性中心结合的大概3bp的双螺旋
活性中心是一个疏水环境
因为2'-OH的空间位阻
螺旋上的脱氧核糖磷酸骨架呈锯齿状或Z字型伸展
螺旋的方向是左手
G的糖苷键是顺式
平坦
极度窄,很深
减少两条链间磷酸基团的斥力
嘌呤-嘧啶相间排列
中和磷酸基团的负电,有利于Z-DNA的形成
在DNA周围创造局部的高盐浓度的微环境
C上的甲基被认为有助于B向Z的转变
有时负超螺旋也能促进Z-DNA的形成
其与B-DNA的交界处会各有一对碱基被从内部挤出来
一般认为与基因的表达调控有关
可能通过释放来自螺旋本身的拓扑学张力而促进同源重组
X射线衍射数据
A=T、C=G、A+G=C+T
如果复制时某些碱基处于烯醇式或亚氨基式,会导致错配
碱基对之间的氢键
对螺旋稳定起决定性作用
碱基对之间在垂直方向上的相互作用所产生的力
包括疏水键和范德华力
嘌呤与嘌呤>嘌呤与嘧啶>嘧啶与嘧啶
碱基的甲基化能提高碱基堆积力
GC堆积力高于AT
碱基对接触面积更大
降低两条链的斥力
取决于DNA环境的离子强度
某区段含有成串的A序列(4-6个A),且相邻的串间隔10个碱基时,容易形成弯曲
受到某些蛋白质的作用
作为DNA修复系统识别的一种信号
有利于压缩比较大的基因组DNA
在人类基因组DNA的复制起始区和转录调控区,有许多反向重复序列
也许能充当某种控制DNA复制和基因转录的开关
在双螺旋的基础上沿大沟引入第三条链
2x嘌呤+1x嘧啶
1x嘌呤+2x嘧啶
第三条链上的C需质子化(低pH)
序列呈镜像重复
仍是A-T、C-G
第三条链与嘌呤链呈平行排列
第三条链与嘌呤链呈反平行排列
碱基堆积力
高盐浓度时对磷酸负电荷的中和
负超螺旋
影响DNA的复制、重组和转录
调控基因表达
DNA双螺旋上的某个碱基离开其配偶,突出在双螺旋之外
参与同源重组的酶需要通过碱基翻转寻找同源的序列
催化碱基修饰的酶需要碱基通过翻转落入其活性中心被化学修饰
参与碱基切除修复的DNA糖苷酶需要受损伤的碱基通过翻转落入其活性中心
DNA分子上的U就是通过这种方式被尿嘧啶-DNA糖苷酶切除
可能导致某些基因发生移框突变
若此时复制,游离在外的环可能被跳过
两个两侧含有两段G四联体(GGGG)重复序列的GCGC序列组成的DNA片段间
端粒是最可能形成四链结构的地方
已知一级结构,可以对其二级结构进行很好的预测
2'-OH的空间位阻导致的
两个氢键
增加了形成局部双螺旋的可能性
最常见的二级结构是发夹结构(茎环结构)
73-94个核苷酸
部分碱基常甲基化
5S
16S
23S
5S
5.8S
18S
28S
使rRNA高度折叠
不同物种的rRNA一级结构并不相似,但二级结构相似度很高
可调控基因表达
最常见的是茎环结构
松弛型
过度缠绕
左手螺旋
不利于复制、重组、转录
可通过DNA拓扑异构酶清除
缠绕不足
右手螺旋
利于复制、重组、转录
在螺旋轴被限制在平面上,双螺旋DNA的一条链以右手的方向环绕螺旋轴的总次数
L=T+W
DNA的双螺旋的总转数
双螺旋的轴环绕超螺旋轴的次数
超螺旋密度
需要RNA伴侣的帮助
探测细胞内特定物质的浓度变化,并作出反应,影响相关基因表达
很多酶的辅酶
TPP浓度高时能结合TPP代谢有关的基因的RNA
参与形成多种核酶和自我剪接的内含子的活性中心
诱导多种病毒在翻译过程中发生核糖体移框
RNA部分充当端粒DNA合成的模板
共轴螺旋
A-小沟模体
核糖拉链
弯曲-转角
金属离子
碱性蛋白质
在一级结构上相距较远的两端区域形成的Watson-Crick碱基对或非Watson-Crick碱基对
碱基堆积力
碱基和主链之间的相互作用
2'-OH也会参与形成氢键
中和RNA链上的负电荷
有效隔开tRNA的两个功能端
无三级结构
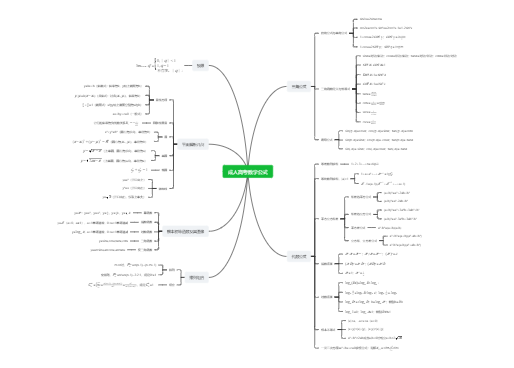
真核细胞的核基因组DNA在细胞核内与组蛋白结合形成的结构
可视为真核生物细胞核染色质的一级结构单位
每一个“珠子”由组蛋白核心和环绕其上的DNA组成
最容易受到DNA酶的水解
去除H1并不会破坏核小体结构
一类较小的碱性蛋白
生理pH下带负电
保守性最低
某些组织中没有H1,用其他组蛋白取代
有利于将核小体包装成更高层次的结构
保守性第三
保守性第三
保守性第二
保守性最高
改变局部的核小体和染色质的结构,可能影响到基因的表达
H1°
H5
N端形成尾
组蛋白折叠
2x(H3+H4)
2x(H2A+H2B)
主要通过静电引力与DNA结合
表面环绕约146bp的DNA双螺旋
DNA序列在核小体选位中起重要作用
可以通过启动远距离DNA序列之间的相互作用而激活基因表达
通过阻碍特定转录因子或调节蛋白与DNA的结合而抑制基因表达
DNA只有在从核小体上解离或部分解盘绕的情况下才可与转录因子或调节蛋白充分接触
乙酰化
甲基化
磷酸化
泛酰化
影响其与DNA的相互作用
对核小体的稳定和DNA的解螺旋起调控作用
古菌的基因组也是共价闭环的DNA
古菌也含有组蛋白,也会形成核小体
HMfA
HMfB
短于真核生物
包被约80bp
无法进行化学修饰
细菌没有组蛋白
形成高度浓缩的拟核或类核的结构
小环被固定在由特定的蛋白质分子形成的基座上
拟核蛋白NAP
DNA拓扑异构酶
转录因子
mRNA
核糖体
识别多种真核细胞与粗面内质网核原核细胞与细胞膜结合的核糖体上合成的蛋白质在N端的信号肽,参与这些蛋白质的共翻译定向和分拣
参与细胞核内mRNA的剪接
参与真核细胞rRNA的后加工
切除真核细胞mRNA分子内的内含子
参与tRNA前体在5'端多余的碱基序列的切除
端粒酶
RNA病毒
Avery的细菌转化实验
Chase的噬菌体实验
充当RNA病毒的遗传物质
核酶
参与蛋白质的生物合成
作为引物,参与DNA复制
参与RNA前体的后加工
参与基因表达的调控
参与蛋白质共翻译定向和分拣
参与X染色体的失活